WHAT IS A SPECIES?
Kurlovich, B.S.
Definitions:
(1) The taxonomic rank, and the most basic unit or category of biological classification.
(2) An individual belonging to a group of organisms (or the entire group itself) having common characteristics and (usually) are capable of mating with one another to produce fertile offspring.
(3) A group of living organisms consisting of similar individuals capable of exchanging genes or interbreeding. The species is the principal natural taxonomic unit, ranking below a genus and denoted by a Latin binomial, e.g., Homo sapiens.
Our further reasoning will be applied to plants. Plant identification connects to the science of taxonomy, or classification. Swedish physician Carolus Linnaeus developed the current taxonomical system in the 18th century, classifying plants based on their relationships to each other and organizing them in a hierarchy beginning with kingdom (a collection of organisms that share basic traits) and descending to species, organisms so closely related that they can produce fertile offspring together. Plants belong to the plant kingdom. Determining which taxonomical groups, or taxa, they belong to after that helps you proceed to identifying a precise species. A species is given a two-part name: the generic name and the specific name (or specific epithet). For example, Lupinus albus L.

Carolus Linnaeus
Two famous Russian botanists made a significant contribution to the study of plant species: Nikolaj I. Vavilov and Vladimir L. Komarov. Ideas of V.L. Komarov (later a supporter of Lysenko) were based on monotypic species, according to which species cannot include a systematic unit of a lower rank (Komarov, 1931; Krassilov, 1989).

Vladimir L. Komarov
The concept of a biological species based on no crossing between species was also widely accepted (Grant 1981, 1984). N.I.Vavilov, however, in a study of several hundred species showed the absence of monotypic species, i.e. species represented by various forms. N.I. Vavilov formulated a polytypic species concept at the 5th INTERNATIONAL BOTANICAL CONGRESS in Cambridge in August, 1930; put forward the ideas of multiformity of species, relativity of taxonomic criteria and the theory of peripheral accumulation of recessive characters. From the evolutionary point of view he considered species as knots in evolutionary chains. Vavilov sought an aphoristic definition of the kind given by the then also well-known botanist V.L. Komarov: a species is a morphological system plus geographic distinctness. Elaborating on this, Vavilov defined a "Linnaean species" as "an isolated complex dynamic morph-physiological system bound in its origin to a certain environment and area" (Vavilov,1931a, p.215).
N.I. Vavilov’s whole attention has been devoted to the intraspecific diversity of separate plants, to the botanical study of the varietal diversity of separate Linnaean species including cultivated plants, and to the study of the geography of varieties in ancient and modern agricultural countries. The investigation of several hundred cultivated species, carried on by a group of scientific workers, according to a strictly defined plan, has led to the conception of a Linnaean species as a definite system.

Nikolaj I.Vavilov
The chief regularity determining the diversity of a Linnaean species, its system, has been called by Vavilov «the Law of homologous series in variation» (Vavilov, 1920). The term "homologous" applies according Vavilov chiefly to the species within the limits of the same genus. In comparing different genera it would be sometimes more to the point to speak of parallelism in analogous variation. In studying the one or the other species, we, as a rule, draw up a system of hereditary variation for all characters, including the characters of the reproductive organs, as well as the vegetative characters and the biological peculiarities. The vast mass of facts has led Vavilov to the conception of a Linnaean species as a system of forms, the building up of which is controlled by the law of homologous series. The conception of a Linnaean species, as a regular system, seemed to Vavilov of great importance for practical purposes connected with the investigation of cultivated plants, as well as for the botanical study of the principal question of the process of evolution. A close study of the evolution process, in the light of Vavilov’s conception of a species, is possible only in considering the Linnaean species as a system. Only the knowledge of the geography of the chief potential or varietal characters (genes) will lead to a real mastery of the species (N.I. Vavilov, 1931a, p.214). He has suggested that the method of differential systematic and differential geography should be used for the study of species. With regard to plants which are of importance in cultivation, this is the only way of mastering the Linnaean species. The study of a species embraces not only morphological, but also physiological characters. Many Linnaean species represent a complex system of ecotypes or climatypes. It is natural that, differentiating in space, and under going the influence of selection, the principal potential of a Linnaean species singles out groups of hereditary forms most adapted to the given environment. Thus, a Linnaean species is according to Vavilov’s conception (1931a, p.215), a separate morph physiological system connected in its genesis with a definite environment and area. In opposition to the rather common notion as to the relativeness of species and the conventional character of their understanding, the great number of concrete facts encountered in studying the cultivated plants and their wild relatives has induced Vavilov to recognize the Linnaean species as actual complexes, actual systems, which exist in Nature and represent important definite links in the evolutional chain, the know ledge of which is very helpful in mastering the multifariousness of the organized world. The concrete material shows that the so-called "Linnaean species, may be very different in regard to their contents. Being more or less separate systems, they manifest themselves in a different compass. The analysis of a great number of Linnaean species by means of the method of differential systematic and differential geography, as well as by the modern methods of genetics and cytology, reveals the great diversity of the species. Thus, there are sometimes bulky systems be![]() longing to one Linnaean species, which might be expediently subdivided into categories or subspecies. Sometimes, on the contrary, a Linnaean species represents a very limited and comparatively small system. The law of homologous series in hereditary variation gives an answer to the question what material should be looked for, while the theory of the centers of origin of cultivated plants provides an answer to the question where it could be found. To study intraspecific diversity and to determine the centers of origin of cultivated plants, Vavilov and his followers used a differential systematic and geographical method of crop studies (Vavilov, 1931a,b) which meant as follows: · differentiation of a genus into species and intraspecific diversity with the help of morphological, hybridylogical, cytological and others methods; determination of the genotypic composition of a species; geographical localization of hereditary forms of a species as well as the centers of their diversity. Such approach supported by the development of different intraspecific classifications helped to accomplish profound and comprehensive analysis of intraspecific and varietal diversity of cultivated plants, and find ways of efficient utilization. In this respect, the International Code of botanical nomenclature fixed such categories as subspecies (subspecies), varieties (varietas), subvarieties (subvarietas) and form (forma).
longing to one Linnaean species, which might be expediently subdivided into categories or subspecies. Sometimes, on the contrary, a Linnaean species represents a very limited and comparatively small system. The law of homologous series in hereditary variation gives an answer to the question what material should be looked for, while the theory of the centers of origin of cultivated plants provides an answer to the question where it could be found. To study intraspecific diversity and to determine the centers of origin of cultivated plants, Vavilov and his followers used a differential systematic and geographical method of crop studies (Vavilov, 1931a,b) which meant as follows: · differentiation of a genus into species and intraspecific diversity with the help of morphological, hybridylogical, cytological and others methods; determination of the genotypic composition of a species; geographical localization of hereditary forms of a species as well as the centers of their diversity. Such approach supported by the development of different intraspecific classifications helped to accomplish profound and comprehensive analysis of intraspecific and varietal diversity of cultivated plants, and find ways of efficient utilization. In this respect, the International Code of botanical nomenclature fixed such categories as subspecies (subspecies), varieties (varietas), subvarieties (subvarietas) and form (forma).
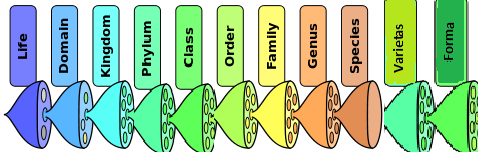
The hierarchy of biological classification's taxonomic ranks. Intermediate minor rankings are not shown!
Besides, Vavilov gave special heed to eco-geographical differentiation of the species into ecotypes, geotypes, concultivars, etc. (Vavilov, 1931b, 1965a; Sinskaja, 1969). The postulate of the species as a complex multilateral phenomenon would urge a researcher to use diverse methods for identifying differences between intraspecific categories. In addition to the main morphological method, the scientists of the Vavilov Institute have widely used anatomic, cytological, caryological, paleobotanical, ontogenetic, biochemical, physiological, geographical, genetic and other methods. Such comprehensive approach is especially efficient when the intraspecific diversity of cultivated lupin forms is concerned. This Vavilov’s differential systematic and geographical method of crop studies is interconnected with other findings of N.I. Vavilov and is based on his law of homologous series in hereditary variation, his theory of the centers of origin (diversity) of cultivated plants, and his concept of the species as a system (Вавилов, 1920, 1926, 1935, 1987b).
All Vavilov’s fundamental ideas interconnected among themselves and represent a complex doctrine about global genetic diversity of cultivated plants. On the basis of this doctrine, the experts of the Vavilov Institute (VIR) have developed intraspecific classifications practically for all leguminous crops. Vavilov invited his friend L.I. Govorov to manage the Leguminous crop department, and he recruited experts in leguminous plants to the staff. Govorov developed intraspecific classification for peas (Govorov 1937), organized breeding work with leguminous crops in the former USSR and created many pea cultivars. Like Vavilov, Govorov had a tragic destiny; he was arrested and disappeared. P.M. Zhukovsky was invited to manage the lupin collection. He developed and published important works on interspecific and intraspecific diversity (Zhukovsky 1929). Research on the genetic resources of lentil was done by E.I. Barulina (Vavilov’s wife). She took part in many expeditions, e.g. to Crimea, Georgia and other regions (Barulina 1930). During the Second World War the department of leguminous crops was managed by N.R. Ivanov, who made a substantial contribution to preserving the collection during the seige of Leningrad. He carried out important research on the genetic resources of kidney bean (Ivanov 1961). After the Second World War, under the direction of Korsakov, ecological and geographical research at VIR was modified and databases on genetic resources were continued. Korsakov also contributed to the clarification of centres of origin of various soya bean species (Korsakov 1971).

N.I.Vavilov Institute of Plant Genetic Resources (VIR)
Intraspecific classifications for peas was developed (Govorov, 1937; Makasheva, 1973, 1979), mung bean (Popova, 1937), soybean (Korsakov, 1971, Teplyakova, 1997), chickpea (Seferova, 1997), and vetch (Stankevich and Rep’ev, 1999). These classifications make it possible to disclose completely the potential of leguminous crops, and they are widely used in breeding practice and plant science. Numerous scientists in many countries continue the traditions and methods of Vavilov. On the field of cultivated crops, especially legumes, ideas of Vavilov have been further developed in Australia (Gladstones, 1974, 1998; Sweetingham, 1986, 1989; Cowling, 1994; Clements and Cowling, 1990, 1991), Finland (Hovinen, 1994; Tigerstedt, 1994), Germany (Diederichsen and Hammer, 1996), Italy (Laghetti et al., 1996; Saccardo, 1996), Poland (Swęcicki W, 1988; Swęcicki W.K., 1996; Kazimierski and Kazimierska, 1992; 1994), Portugal (Mota, 1984; Gusmão, 1988 ; Gusmão et al., 1996; Tavares de Sousa et al., 1992; Neves Martins; 1994; Dordio, 1996), Sweden (Blixt, 1970, 1996), UK (Polhill, 1976, Bisby, 1981; Ambrose, 1996) and many others.
We have also reviewed the system of genus Lupinus L. using Vavilov’s concept of the species as a system, and applied his differential systematic and geographical method in our studies. On this topic before us also worked some Russian scientists (Maissurjan and Atabekova, 1974). On the basis of Vavilov’s concept we offer intraspecific and eco-geographical classifications for three cultivated annual species of lupin (L. albus L., L. luteus L. and L. angustifolius L.). This approach provided a possibility to perform a targeted search of genetic resources of lupins to solve problems of their evolution and selection. It enabled us not only to cast light on the diversity of lupin forms, but also to reveal a series of regularities in their variation depending on the degree of cultivation, geographic environments and soil conditions. Such outcome, in its turn, helped in finding and recommending valuable initial material for breeding. N.I. Vavilov laid a foundation of the lupin collection in VIR. Valuable accessions of white lupin (Lupinus albus L.) were collected by Vavilov during his trip to the Mediterranean in 1926. In Palestine, he managed to find very early, thermally neutral and small-seeded forms relating to the Jordanian ecotype. In particular, the duration of growing period of the sample Tel Karam (k-290) in the conditions of the Ukraine was only 105 days. From Sudan Vavilov brought, on the contrary, very late, but highly productive and large-seeded forms belonging to the Sudanese ecotype (k-486, k-495). Of special value as sources for lupin breeding in Russia, Poland and the Ukraine were samples of the Jordanian ecotype from Palestine. Their hybridization with samples of the Georgian ecotype (Kurlovich, 1996), and also their involvement in combinations with application of mutagenesis, allowed V.I. Golovchenko to create early and high productive cultivars of white lupin, Kievsky mutant, Horizont and Druzba, suitable for the condition of Russia and the Ukraine (Головченко et al, 1984).

B.S. Kurlovich
Not only did Vavilov collect and organize studying of the assembled plant materials, but also he constantly strove to improve methods of these studies. He took the initiative, in particular, to develop a simpler and more widely accessible method of determining alkaloids in lupin. This effort of his was inspired by the fact that the technique of detecting and determination of lupin forms with low alkaloid content elaborated in 1928-1929 by v. Sengbusch in Germany was kept secret, since sweet strains of lupin were sold to a private firm. As a result, under the leadership of Prof. N.N. Ivanov, M.I. Smirnova and other experts of VIR worked out an efficient rapid method for determination of alkaloids by means of different chemical reagents. It was immediately published for the first time in the world with the Introduction by N.I Vavilov (Иванов et al., 1932). This publication, together with the know-how of German scientists, launched the breeding work with fodder (sweet) low-alkaloid lupin on a worldwide scale.
Each form of lupin can be assessed as an object fitting in with a number of consecutive taxa. We recognize subspecies as an isolated group of individual plants within a population of a species in relation to lupins. They occupy certain part of the area of a species, constitute together a mobile system, are able to cross among themselves and with plants growing in other parts of the area of this species, produce prolific progeny, possess distinctive morphological and inheritable characters in vegetative and generative organs with the uniform genetic base, and incorporate transient forms.
Allelism and character complementarities tests have shown that in lupins the color of seed is correlated with the color of the corolla. This linkage reflects the stability of genetic system, which corresponds to the rank of varieties.
A good diagnostic character is the color of vegetative parts, and the absence or presence of anthocyan, in particular. Being less stable, it could be used in identifying subvarieties.
Considerable practical interest for breeders may be generated by the plants with determinate branching, fascicular stem and other characters of breeding value. Such forms are theoretically possible in all the varieties and subvarieties. Therefore, it seems justified to regard them in the rank of form (f.). Intra - specific classification of white lupin (Lupinus albus L.) developed by us looks as follows:
The scheme of classification of Lupinus albus L.
I. Subsp. graecus (Boiss. et Spun.) Franko et Silva
2. Subsp. termis (Forsk.) Ponert.
I. Var. abissinicus Libk.
2. Var. subroseus Libk.
3. Subsp. albus L.
3. Var. albus
4. Var.vavilovii (Atab.) Kurl. et Stankev.
5. Var. vulgaris Libk.
I. f. libkindae Kurl. et Stankev.
Lupinus albus is widely spread as wild plants and Cultivated over all the Mediterranean region and many Countries of the World! The ancient culture of white lupin under the local name «hanchcoly» until recently existed in Western Georgia (Kurlovich, 1996a). The closest described neighbors of this species were: L. graecus and L. termis . The first is the wild plant on Balkan Peninsula; the second is grown in Egypt, Libya, Sudan, Ethiopia, Israel, Palestine and Syria. Information accumulated by scientists made it possible to revise and integrate the volumes of these species.
All three species differ only in the colour and size of flowers and seeds (quantitative characters). Cultivated forms of white lupin were introduced in agricultural practice long time ago, before the ancient times, on the place of present-time Greece. To the south (Egypt, Libya, Palestine) mainly the forms with white seeds and pink-and-blue or light-pink flowers (L. termis) were spread, while westwards (Apennine Peninsula and farther) mostly the forms with white seeds and grayish-blue or white seeds (L.albus) dispersed. White lupin (L.albus) is still grown in Greece, where wild L.graecus is also spread. But, The absence of distinct specific characters makes it possible to consider all three species harbored within the limits of one species of Linnaeus (Lupinus albus). But the existence of geographical and physiological differentiation is sufficient to recognize them as having the rank of subspecies in L.albus L: subspecies. albus, subspecies graecus (Boiss. et Sprun.) Franco et Silva, and subspecies termis (Forsk.) Ponert (Kurlovich, 1990, 1998; Kurlovich et al. 1995).
Using the developed criteria of intraspecific taxa, VIR’s blue lupin collection (L. angustifolius L.) was screened to identify 13 varieties distinctly differing from each other in the colour of seeds and the corolla. Variations in the colour of cotyledons, vegetative parts and carina have been used for identification of 12 subvarieties. Besides, the plants with determined branching and fascicular stems are described as 8 separate forms. Variability of characters at yellow lupin (Lupinus luteus L.) is less expressed than in L. angustifolius; however, a homologous series can be modelled on the colour of seeds, which is more or less similar to L. angustifolius. Dominating colouring of flowers is yellow or, less frequently, lemon-yellow, orange or whitish. Using combinations of such characters as the colour of the corolla, the carina’s edge, vegetative organs and seeds, 18 varieties, 4 subvarieties and 6 forms have been identified (Kurlovich, 1990, 1998, 2002, 2015).
Effective utilization of plant genetic resources in plant breeding requires detailed information about different biological, physiological, biochemical and other properties of accessions.
Eco-geographic classification reflects the regularities of variability of these properties, based on the specific nature of geographic, historical, ecological and agronomic conditions. It cannot replace the botanical classification of the species, which requires clear distinction of morphological characters observed in the herbarium materials, but supplements it. According Vavilov, the species is: "... a flexible isolated complex morpho-physiological system linked in its genesis to a certain environment and area of distribution". Therefore, the species should be studied by morphological, physiological, biochemical, genetic and other methods.
We were guided in our research by Vavilov’s ideas, and we arranged the genetic resources of lupins into: geotypes, eco-geographic groups of ecotypes, separate ecotypes and concultivars (varietal types).
The accessions of each lupin species adapted to a definite and sufficiently vast habitat or geographic area of cultivation were grouped into geotypes, typical of the species with wide areas of distribution. Prof. Agaev ( 1987 ) defined the term “geotype” as "morphologically weakly differentiated but genetically determinate race, that appeared in the composition of species as a result of their settlement and evolution".
Geotypes were divided into separate ecotypes or eco-geographic groups of ecotypes (when it was difficult to select separate ecotypes).
Ecotypes are detected among plants adapted to specific niches in the wild conditions, e.g. in mountains, on littoral, close to villages or roads) or in agriculture (e.g. different local forms). "An ecotype is a group of biotypes within one Linnaean species united by a series of constant hereditary characters and adapted to certain conditions of their habitat. It is quite natural that differing in space and submitting to the effect of selection, the basic potential of the Linnaean species is to develop and isolate the groups of hereditary forms most closely corresponding to the given environments..."(Vavilov 1965, p.245).
Breeding varieties were classified into concultivars (varietal types).
We have grouped the gene pool of white lupin available now in the VIR collection into 11 geotypes and agrogeotypes, 4 ecogeographic groups of ecotypes, 17 separate ecotypes and 10 concultivares. Ecogeographic research has shown the ranges of changeability in different descriptors of white lupin forms. Duration of growing period under spring sowing varies from 106 to 180 days, seed mass per plant changes from 2.2 to 40 g, green mass yield per from 9 to 250 g, protein content in seed from 35.0 to 53.7%, and oil content from 6.2 to 12.0% . The diversity of narrow-leafed lupin is grouped into 12 geotypes and agrogeotypes, 16 separate ecotypes, 3 ecogeographic groups of ecotypes, and 13 concultivars. The diversity of yellow lupin is classified into 8 geotypes, 10 ecotypes, 2 eco-geographic groups of ecotypes, and 10 concultivars (Kurlovich et al. 1995; Kurlovich, 1998, 2002, 2015). The results indicate that the lupin samples of Iberian geotype were the most efficient in breeding for resistance to disease or for fresh green mass. Specimens from the Balkan-Asian and Palestinian geotypes are more efficient in creating cultivars for grain yield, breeding for earlyness, large seed, drought resistance and increased oil content. Wild-growing forms of all the above lupin species undoubtedly bear numerous valuable characters: small seed, drought tolerance, resistance to low temperatures and diseases. Among the forms most adapted for cultivation and breeding in Russia are those that have escaped from domestication, or turned wild, as well as local varieties from the Mediterranean countries.
Conclusions
My colleagues and I are supporters of the concept of Vavilov about a Linnaean species as a definite system. N.I.Vavilov showed the absence of monotypic species, i.e. species represented by various forms. Vavilov formulated a polytypic species concept at the 5th INTERNATIONAL BOTANICAL CONGRESS in Cambridge in August, 1930; put forward the ideas of multiformity of species, relativity of taxonomic criteria and the theory of peripheral accumulation of recessive characters. From the evolutionary point of view he considered species as knots in evolutionary chains. Vavilov sought an aphoristic definition of the kind given by the then also well-known botanist V.L. Komarov: a species is a morphological system plus geographic distinctness. Elaborating on this, Vavilov defined a "Linnaean species" as "an isolated complex dynamic morph-physiological system bound in its origin to a certain environment and area" (Vavilov,1931, p.215). This led to the understanding of the Linnaean species concept, an integral entity consisting of closely interlinked components where the whole and the parts are merged (Vavilov 1965a; Agaev 1987; Korovona 1987). We also have tried to complement and develop the Vavilov’s concept in relation to leguminous plants. Vavilov’s concept of the species as a system have enabled as to find out new regularities in the variability of characters in lupins depending on genetic features of the species, and also on ecological and agronomic conditions of their growth. This approach allowed to develop a more detailed intraspecific taxonomic and eco-geographical classification, and proposed efficient ways of lupins genetic resources utilization in breeding.
Vavilov was the first to recognize the necessity for intensive plant collecting and preservation. He was a highly qualified collector of plant genetic resources and manager of collecting missions, and an author of many theoretical and practical ideas in the field of the global genetic diversity of cultivated plants. A major contribution of Vavilov was his ability to translate his rapidly growing scientific knowledge in genetic resources into economic use. Vavilov’s plant collecting expeditions served as the basis for the leguminous crop collection in VIR, which provided the initial materials for over 75% of new grain legume cultivars created in Russia and other countries of the former USSR (Kurlovich 1996b, 2002).
It is very difficult to do justice to Vavilov in this short article. The scope of Vavilov’s interests was extraordinarily wide and included practically all agricultural crops and disciplines of plant science. The memory of Vavilov has been preserved through his collections of plant genetic resources, ideas, books and followers. These are an important legacy of N.I. Vavilov, and were recently documented posthumously in his book ‘Five Continents’ (Vavilov 1962, 1987a, 1997).
Acknowledgements
We wish to express our gratitude to Prof. M.G. Agaev and Drs A.K. Stankevich and O.N. Korovina for the help, valuable council and participation in the development of intraspecific classifications for leguminous crops.
References
Agaev, M.G. 1987. Vavilov’s conception of species and it development. Genetica XXIII(11):1949-1960.
Ambrose, M. 1996. Grain legume collections and activities in the UK. Pp.100-102 in Report of a Working Group on Grain Legumes (T. Gass, M. Ambrose, J. Le Guen, A. Hadjichristodoulou and S. Blixt, compilers). ECP/GR-Grain Legumes Network, IPGRI, Rome, Italy.
Barulina, E.I. 1930. Lentil in USSR and other countries. Bull. Appl. Bot. Genet. Plant Breed. (Leningrad), Supplement 40:1-319.
Bisby, F.A. 1981. Genisteae. Pp. 409-425 in Advances in Legume Systematics, Part 1 (R.H. Polhill and P.H. Raven eds.). Royal Botanic Gardens, Kew, UK.
Blixt, S. 1970. Studies of induced mutations in peas. XXVI. Genetically conditioned differences in radiation sensitivity. Agr. Hort. Genet., Bd. 28:55-116.
Blixt, S. 1996. Status of legume collections in the Nordic countries. Pp. 61-62 in Report of a Working Group on Grain Legumes (T. Gass, M. Ambrose, J. Le Guen, A. Hadjichristodoulou and S. Blixt, comps). ECP/GR-Grain Legumes Network, IPGRI, Rome, Italy.
Cowling, W.A. 1994. Use of lupin genetic resources in Australia. Pp. 9-18, in Advances in Lupin Research (J.M. Neves Martins and M.L. Beirao da Costa, eds.). Proceeding of the 7th International Lupin Conference, Evora, Portugal. ISA Press, Lisbon, Portugal.
Diederichsen, A. and K. Hammer. 1996. Status of plant genetic resources and research on grain legumes in Germany. Pp.45-50 in Report of a Working Group on Grain Legumes (T. Gass, M. Ambrose, J. Le Guen, A. Hadjichristodoulou and S. Blixt, comps). ECP/GR-Grain Legumes Network, IPGRI, Rome, Italy.
Gladstones, J.S. 1974. Lupin of the Mediterranean region and Africa. W. Aust. Dept. Agr. Tech. Bull. 26. 48 p.
Gladstones, J.S. 1998. Distribution, origin, taxonomy, history and importance. Pp. 1-39 in Lupin as Crop Plants. Biology, Production and Utilization (J.S. Gladstones, C. A. Atkins and J. Hamblin, eds.). Cambridge University Press, Cambridge, UK.
Golovchenko, V.I., N.S. Kucherenko, N.G. Lesunova and A.S. Porokonnyij. 1984. Genefund and the result of breeding fodder lupine. Res. Bull., N.I. Vavilov Inst. Plant Ind., Leningrad, USSR 139:7-12.
Govorov, L.I. 1937. Pea. Pp. 229-336 in Flora of Cultivated Plants. State Printing Office, Moscow and Leningrad, USSR.
Grant, V. 1981. Plant Speciation. Columbia University Press, New York, USA. 528 pp.
Grant, V. 1984. Plant Speciation. Transfer with English. Publishing House – World, Moscow, USSR. 528 pp.
Hill, J., H.C. Becker and P.M.A. Tigerstedt. 1998. Quantitative and Ecological Aspects of Plant Breeding. Chapman & Hall, London, UK. 275 p.
Hovinen, S. 1994. Tiina – uusi herneen yleislajike. Koetoiminta ja Kдytдntц 51:35.
Ivanov, N.N., M.I., Smirnova, N.I. Sharapov et al. 1932. Problem of alkaloid-free lupin. Exhibit 54 to Bull. Appl. Bot. Genet. Plant Breed. 63 pp.
Ivanov, N.R. 1961. Kidney bean. State Printing Office, Moscow and Leningrad, USSR. 280 pp.
Kazimierski, T. and E.M. Kazimierska. 1992. Natutalne tetraploidy lubinu bialego (Lupinus albus L.). Materialy konferencji “Genetika 2000”. Krakуw, Poland.
Kazimierski, T. and E.M. Kazimierska. 1994. The course of meiosis and microsporogenesis in dihaploid plants of white lupin (L. albus L.). Genet. Pol. 35(3):153-160.
Komarov, V.L. 1931. Origin of cultivated plants. State Printing Office, Moscow and Leningrad, USSR. 200 pp. Komarov, V.L. 1944. The doctrine about a species at plants. Leningrad, USSR. 285 pp.
Korovina, O.N. 1987. N.I. Vavilov on the primary centres of cultivated plant origin. Bot. Journal, St. Petersburg 6:721-729.
Korsakov, N.I. 1971. Geographical seats of soybean formation. Bull. Appl. Bot. Genet. Plant Breed. 45:189-193.
Krassilov, V.A. 1989. Vavilov's species concept and evolution of variation. Evolutionary Theory, 9: 37-44.
Kurlovich, B.S. 1989. On the centers of species formation of the genus Lupinus L. Bull. VIR 193:20-24.
Kurlovich, B.S. and A.K. Stankevich. 1990. Intraspecific diversity of three annual lupine species (Lupinus L.). Bull. Appl. Bot. Genet. Plant Breed. 135:19-34.
Kurlovich, B.S., S.I. Rep’ev, L.G. Shchelko, V.I. Budanova, M.V. Petrova, T.V. Buravtseva, A.K. Stankevich, L.T. Kartuzova, T.G. Alexandrova, T.E. Teplyakova and L.K. Malysh. 1995. Theoretical basis of plant breeding. Vol. III. The gene bank and breeding of grain legumes (lupine, vetch, soya and bean). VIR, St. Petersburg, Russia. 438 p.
Kurlovich, B.S. 1996a. Georgian ecotype of white lupine (Lupinus albus L.). Plant Genetic Resources Newsletter, Rome, 108:66-68.
Kurlovich, B.S. 1996b. Genetic resources of grain legume crops in N.I. Vavilov All-Russian Research Institute of Plant Industry. Pp.77-81 in: Report of a Working Group on Grain Legumes (T. Gass, M. Ambrose, J. Le Guen, A. Hadjichristodoulou and S. Blixt, comps). ECP/GR-Grain Legumes Network, IPGRI, Rome, Italy.
Kurlovich, B.S. 1998. Species and intraspecific diversity of white blue and yellow lupins. Plant Genet. Resources Newsletter. 115:1-10.
Kurlovich, B.S. 2002. LUPINS: Geography, Classifications, Genetic Resources and Breeding.// St. Petersburg, Publishing house «Intan», 468p.
Kurlovich, B.S. 2015. Diversity of Lupinus albus L., Lupinus angustifolius L. and Lupinus luteus L. P.142 in: Developing lupin into a major and sustainable food and feed source (Jessica Capraro et al. eds.). Proceedings of the 15th International Lupin Conference, Milan, Italy, 21-26 Lune 2015.
Maissurjan, N.A. and A.I. Atabekova. 1974. Lupine. Kolos, Leningrad, USSR. 463 p.
Makasheva, R. Kh. 1973. Pea. Kolos, Leningrad, USSR. 311 p.
Makasheva, R. Kh. 1979. Pea. Flora of cultivated plants. V. IV, part 1. Kolos, Leningrad, USSR. 322 p.
Mota, M. 1984. Lupin germplasm resources in Portugal. p. 8 in Abstracts of the IIIrd International Lupin Conference, La Rochelle, France.
Neves Martins, J.M. 1994. Numerical taxonomy on the study of Lupinus albus accessions. Pp. 84-89 in Advances in Lupin Research: Proceedings of the 7th International Lupin Conference, Evora, Portugal, 1993 (J.M. Neves Martins and L.M. Berдo da Costa, eds.). ISA Press, Lisbon, Portugal.
Pavlova, A.M. 1952. Varieties of mung bean for new regions of irrigation. Bull. Appl. Bot. Genet. Plant Breed. 29(3):67-70.
Polhill, R.M. 1976. Genisteae (Adans.) Benth. and related tribes (Leguminosae). Bot. Systematics 1:143-368. Popova, G.M. 1937. Phaseolus aureus (Roxb.). Piper – Mung pea. Pp. 573-601 in Flora of cultivated plants. VIR, Leningrad, USSR.
Rep’ev, S.I. 1988. Principles governing selection of parental pairs in the transgressive breeding of common vetch. Bull. Appl. Bot. Genet. Plant Breed. 117:69-76.
Rep’ev, S.I. and V.N. Barulin. 1998. Human Being: Information, Systems of Interaction, Heredity. Theory and Practice. Intan, St. Petersburg, Russia. 116 pp. (in Russian).
Saccardo, F. 1996. Status of grain legume collection in Italy. Pp. 53-56 in Report of a Working Group on Grain Legumes (T. Gass, M. Ambrose, J. Le Guen, A. Hadjichristodoulou and S. Blixt, comps). ECP/GR-Grain Legumes Network, IPGRI, Rome, Italy.
Seferova, I.V. 1997. Varieties of chickpea – Cicer arietinum L. Bull. Appl. Bot. Genet. Plant Breed. 152:9-18.
Sinskaja, E.N. 1969. The historical geography of flora of cultivated plants. Leningrad, USSR. 480 p.
Stankevich A.K. and S.I. Rep’ev, 1999. Vetch. Flora of cultivated plants, 490 p.
Swęcicki, W. 1988. Lupin gene resources in the Old World. Pp.2-11 in Proceedings of the 5th International Lupin Conference (T. Twardowsky, ed.). PWRiL, Poznan, Poland.
Sweetingham, M.W. 1986. Research into lupin root diseases. West. Aust. J. Agr. 27:49-52.
Sweetingham, M.W. 1989. Fungi associated with root and hypocotyl diseases of seedling lupin. Research into lupin root diseases. West. Aust. J. Agr. 40:781-789.
Tavares de Sousa, M.M., I. M. Duarte and M.G. Pereira. 1992. Potential production of winter chickpea in southern Portugal. In: Proceedings of the International Food Legume Research Conference II, Cairo, Egypt, 12-16 April.
Teplyakova, T.E. 1997. Nomenclature analysis of the basis taxonomic systems in soya (Glycine max (L.) Merr.). Bull. Appl. Bot. Genet. Plant Breed. 152:111-118.
Tigerstedt, P.M.A. 1994. Adaptation, variation and selection in marginal areas. Pp. 13-19 in Breeding Fodder Crops for Marginal conditions (O.A. Rongli, E. Solberg and I. Schjelderup, eds.). Kluwer Academic Publishers, Dordrecht, The Netherlands.
Vavilov, N.I. 1920. The Law of Homologous Series in Hereditary Variation. Pp. 3-20 in Proceedings of the III All-Russian Plant Breeding Congress, Saratov, Russia.
Vavilov, N.I. 1926. Centres of origin of cultivated plants. Bull. Appl. Bot. Genet. Plant Breed. 16(2). 248 p.
Vavilov, N. I. 1931a. The Linnean species as a system / N. I. Vavilov (prof.) = [Линнеевский вид как система / Н. И. Вавилов] // Fifth International botanical congrtss, (Cambridge, 16–23 august, 1930) : report of proceedings. – Cambridge : University press, 1931. – P. 213–216.
Vavilov, N.I. 1931b. Linnaeus species as a system. Bull. Appl. Bot. Genet. Plant Breed. 26(3):109-134.
Vavilov, N.I. 1935. Theoretical basis of plant breeding. VIR, St. Petersburg, USSR.
Vavilov, N.I. 1940. The doctrine about an origin of cultural plants after Darwin. Sov. Sci. 2:55-75.
Vavilov, N.I. 1962. Five Continents. State Publishing House of the Geographical Literature, Moscow, USSR. 225 p.
Vavilov, N.I. 1965a. Linnaeus species as a system. pp. 223-250 in The selected works. 5 vols. State Printing Office, Moscow and Leningrad, USSR.
Vavilov, N.I. 1965b. Origin and geography of cultural plants. p. 179 in The selected works. 5 vols. State Printing Office, Moscow and Leningrad, USSR.
Vavilov, N.I. 1987a. Five Continents. Science Publishing House, Leningrad, USSR. 213 pp.
Vavilov, N.I. 1987b. Origin and geography of cultivated plants. VIR, St. Petersburg, USSR.
Vavilov, N.I. 1991. Near the Pamirs (Darvaz, Roshan, Shugnan). Agricultural essay. Bull. Appl. Bot. Genet. Plant Breed. 140:1-12.
Vavilov, N.I. 1997. Five Continents (L.E. Rodin, S. Reznik and P. Stapleton, eds.). IPGRI, Rome, Italy. 198 p.
Zhukovsky, P.M. 1929. A contribution to the knowledge of genus Lupinus Tourn. Bull. Appl. Bot. Genet. Plant Breed. 21(1):16-294.
Zhukovsky, P.M. 1971. Cultivated plants and their relatives. Kolos, Leningrad, USSR. 751 p.
27.02. 2016