Objective regularities in the variability of chatacters

















The genus Lupinus L. numbering some hundreds species of different age and origin is a perfect object for studying the genetic bases of evolution, plant speciation and phylogenesis of plants. A definite combination of the colours of seed, vegetative organs, inflorescences and the carina’s edge s in various lupin species from both hemispheres, as well as complete parallelism in the variability of morphological and biological characters, speaks about their doubtless genetic affinity. The use of the law of homologous series in hereditary variation enabled us to identify and describe several new forms of different lupin species, so far unknown, and to develop intraspecific classification for them. Lupin species more or less closely related to each other are characterized by similar series of variation with such regularity that, knowing a succession of varieties in one species, one can forecast the existence of similar forms in other species. The similarity is more complete in the series of variation, as the species are more closely allied in the general system.
The law of homologous series in hereditary variation gives the answer to the question what material should be looked for, while the theory of the centres of origin of cultivated plants responds to the question where it should be looked for. Vavilov considered the Mediterranean region and mountain areas of Mexico, Peru and others American countries as the centres of origin for this genus (Vavilov, 1926). Generalization based on the differential systematic-geographical method of crop studies provided more precise definition of the centres of formation and origin (diversity) of some lupin species (L. albus L., L. luteus L. and L. angustifolius L.). According to our data (Kurlovich, 1998), the centre of formation of wild white lupin (L. albus L.) and the primary centre of origin (diversity) of its initial cultivated forms is the Balkans where an exceptionally wide diversity of wild and local forms as well as those that turned wild is concentrated. All three subspecies of white lupin (subsp. graecus Franko et Silva, subsp. termis Ponert., subsp. albus) occur in the Balkans. It is mainly this region where wild forms with dotted dark-brown seeds and dark-blue flowers are found (subsp. graecus). Additional confirmation is the Greek name of white lupin: “thermos” (hot). The centres of diversity of cultivated white lupin also include the Apennines and Egypt where its cultivated forms originated in the ancient times. Moreover, in the ancient Egypt forms with pink-and-blue or light pink flowers were widespread (subsp. termis), while on the Apennines forms with greyish-and-light-blue or white flowers were found (subsp. albus).
Existence on the Pyrenees of two close species (L. luteus L. and L. hispanicus Boiss.et Reut.) with the same number of chromosomes (2n=52), wide diversity of wild and cultivated forms of yellow lupin (L. luteus L.), and long historical period of their growing give the reasons to suppose that the Pyrenees harboured the centre of formation of wild yellow lupin forms and the centre of diversity of its cultivated plants (Kurlovich, 1998). For similar reasons, we also regard the Pyrenees as the centre of diversity for narrow-leafed lupin (L. angustifolius L.).
As a result of further agricultural development of lupins, their area of distribution gradually increased. Accordingly, secondary macro- and micro-centres of diversity of cultivated lupin forms emerged on different continents. These secondary centres correspond to the numerous geotypes and ecotypes described by us. In our opinion (Kurlovich, 1998), the present-day secondary centres of diversity of cultivated white lupin (L. albus L.) include France, Germany, Poland, Belarus, Russia and Chile. Secondary centres of diversity of blue lupin (L. angustifolius L.) are, first of all, in Australia, then in Poland and Belarus. To the secondary centres of diversity of yellow lupin (L. luteus L.) Germany, Poland, Belarus, Russia, and Ukraine may be attributed.
Our eco-geographical researches fully confirm Vavilov’s concept about primary localization of dominant genes, first of all, in the primary centres of origin (diversity) and localization of recessive genes at the edges of the species’ distribution area and in the secondary centres of diversity. The forms of lupin occurring within the primary centres more often possess dominant characters, basically typical of wild forms, i.e. strongly pigmented flowers, seed and vegetative organs, dehiscent pods, and monopodial branching. Domesticated forms cultivated mainly in the secondary centres typically have larger white seed, non-dehiscent pods and limited branching (recessive attributes).
Vavilov’s differential systematic-geographical method of crop studies provided a possibility to perform targeted search for valuable breeding materials in various regions and, at the same time, solve the problems of phylogenesis, taxonomy and evolution. This enabled us not only to disclose the diversity of forms, but also to identify a series of regularities in their variation depending on the degree of cultivation and on the geographic, environmental and soil conditions.
Annual domesticated Mediterranean species of lupin, now widely cultivated in different countries (L. albus L., L. luteus L., L. angustifolius L. and others) belong to the group of cultivated plants which maintain close connections with their wild-growing ancestors. Plants of this type only partially transitioned into domesticated state, while in the other part they retain a structure common for the wild flora of the Mediterranean and consist of various ecotypes.
Charles Darwin came to the conclusion that "... Each species was formed in one area and was settled so far, as it was possible ". The species of lupin also occupied their potential areas, and thus adapted to new habitats as a result of their development and domestication.
The Mediterranean region, as the centre of formation of wild-growing lupins, was at the same time one of the first places of their domestication. White lupin was domesticated in the antiquity in Greece (Gladstones, 1974). Yellow and narrow-leafed lupins were cultivated in Spain and Portugal for green manure (Maissurjan and Atabiekova, 1974). Some domesticated plants in these places subsequently turned wild again for different reasons, but continued to grow on the edges of fields and roads. The publication of Klinkowski (1938, 1939) testifies to it.
Domesticated lupins were distributed from the Mediterranean region into other countries: in the area of influence (Sinskaja, 1969). The areas of influence for the ancient Mediterranean were Middle and Northern Europe (Sinskaja, 1969). Lupins were first introduced in these countries, and later underwent certain changes caused by different environmental conditions. Owing to these circumstances, Mediterranean and European countries possess extremely diverse genetic resources of lupin, significant part of which have been accumulated in the collection of the Vavilov Institute (VIR) through plant collecting efforts and inter-country germplasm exchange.
It is possible to identify the following basic groups representing many lupin species:
- Wild ancestors of cultivated plants.
- Local forms cultivated in different historical periods.
- Plant populations turned wild.
- Breeding sources, gene donors and different mutants created and collected by man.
- Modern commercial cultivars and varieties.
All these five groups are represented by an extremely rich diversity, and incorporate morphological, ecological and geographical distinctions promoted by both natural and anthropogenic factors. Wild plants evolved in various natural conditions, therefore their characters vary depending on the region, soil and climate. Local forms cultivated in different historical periods underwent a new cultural step of evolution. Transition of a part of plants into the wild state also produced an effect on their biological features. It was also one of the stages of their evolution. Our researches have shown that plant populations which have turned wild ripen more regularly, have larger seed, and are more resistant to Fusarium and other diseases as compared with the proper wild forms.
Breeding sources, gene donors and different mutants were developed by man for definite breeding purposes. Modern improved cultivars have passed one more stage of evolution. However, they also differ among themselves in their adaptation to certain environments of cultivation.
Great polymorphism of morphological traits is also revealed in all species of lupin. For example, 4 main colours of the flower corolla and 8 types of seed colour were identified for narrow-leafed lupin, respectively 4 and 9 colour types for yellow lupin, and 5 and 6 types for white lupin. Thus, a certain interrelation between these characters was found in different forms. Such regularities were taken as a basis for the development of interspecific classifications.
It is well known that the species of lupin are differentiated into regional geotypes and local ecotypes over an extensive territory (Kurlovich, 1998). They may differ among themselves in their general structure, plant height, number of stalks and branches, leafiness, length of the period of vegetation, biochemical features, disease susceptibility, frost and drought resistance, and yield parameters. Finally, a species becomes a complex population of ecotypes, or an ecosystem. Differences between ecotypes have genetic nature. All geotypes and ecotypes were developed in the process of interaction between genetic recombination, natural selection and environmental effect.
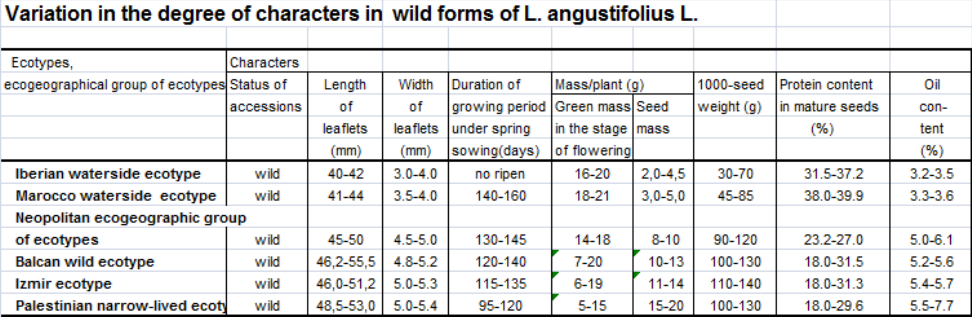
All the studied species of lupin displayed similar ecotypes within the limits of different geotypes with variability governed by Vavilov`s law of homologous series in variation (Vavilov, 1920) and the law of spiral series (Sinskaja, 1969). Thus, different geotypes of the Mediterranean area comprised large-seeded and small-seeded ecotypes, and L. angustifolius L. included ecotypes with very narrow leaflets and also with broader ones. However, there are no ecotypes with absolutely similar features. Geographic regularities were identified in the variability of a number of characters and properties in similar ecotypes. Moving from the west of the Mediterranean region to the east showed an increase in seed yield, seed weight and oil content in seed for all studied species.
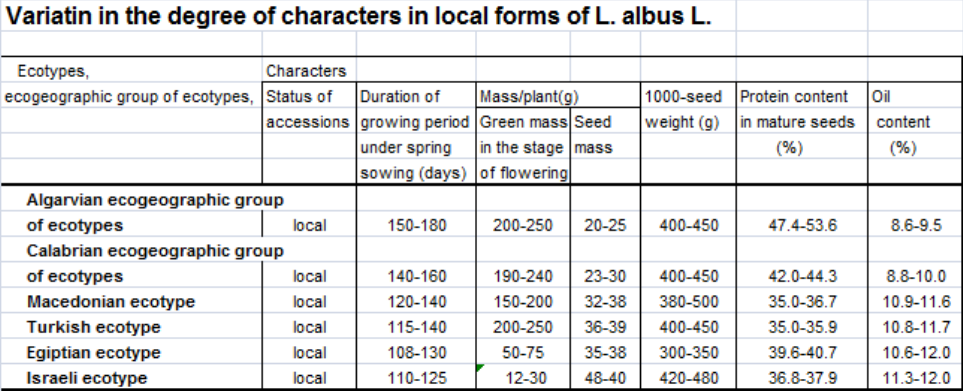
Besides, an increase in the length and width of leaflets was observed in narrow-leafed lupin. Accordingly, the duration of the growing period, green weight of plants, and protein content in mature seeds were decreasing. Moreover, we have found a negative correlation between protein and oil content.
The conducted studies and generalizations helped to formulate answers to the most important questions of plant introduction and breeding, i.e. what kind of genetic diversity should be collected and where, as well as what should be utilized and for what purpose. For instance, lupin accessions of the Iberian geotype proved to be most efficient in breeding for disease resistance or increased green matter yield. Samples of the Balkan-Asian and Palestinian geotypes are more promising for breeding cultivars with higher grain yield, early maturity, larger seed, drought resistance and increased content of oil. Wild-growing forms of all lupin species undoubtedly possess numerous valuable characters, such as small seed, and resistance to drought, low temperatures and diseases. However, their cultivation in Nordic conditions would encounter considerable difficulties, while introduction of their positive properties into breeding programs requires a lot of time. More adapted to cultivation are the forms previously cultivated but later turned wild, and local varieties from Mediterranean countries. The most efficient potential breeding sources of commercially valuable characters (resistance to anthracnose, stress resistance, high seed quality) have been identified among these accessions for the conditions of Russia. Many commercial cultivars from the countries with advanced lupin breeding are also extremely promising for national breeding programs.

Scientific delegation from Australia inspects the collection of lupins at the Kiev experimental site of VIR, 1986.
From left to right: Dr. J.S.Gladstones, translator from Russian into English, Drs. V.I. Golovchenko, W.A. Cowling and M.W.Sweetingham
Breeding programs in lupin-growing countries utilize the most diverse breeding sources and methods mainly because they have advantageous geographical setting. The countries of the Mediterranean region, Australia, New Zealand, Chile and the U.S. have favourable conditions for acclimatization of the great diversity of Mediterranean forms, particularly from the west coastal area of the Mediterranean. Russia and other countries of the former USSR as well as Nordic countries, Poland and Germany are situated in more northern latitudes, and therefore have limited possibilities to use wild Mediterranean forms. Many of them may be cultivated only in glasshouses or artificial climate rooms, although some of them are suitable for autumn sowing in the environments of the Mediterranean and Transcaucasia. Genetic resources from the east coastal area of the Mediterranean have wider application in Russia, because they demonstrate a greater scope of modification variability, have early-ripening habit, and produce larger seed. Besides, in the past years plant breeders in Russia and Poland have also widely used induced mutagenesis. However, the best results might be attained with an optimum combination of mutagenesis and hybridization for different forms of various eco-geographical origin.